
Развитие нервной системы
Развитие нервной системы
Нервные клетки приобретают свои уникальные свойства и образуют четко организованные и удивительно точные синаптические связи во время развития под влиянием генетических факторов и факторов среды. Такими факторами являются: происхождение клеток; индукционные и трофические взаимодействия между клетками; метки, благодаря которым осуществляются миграция и рост аксонов; специфические маркеры, при помощи которых клетки узнают друг друга, а также постоянная реорганизация связей в зависимости от активности клетки.
Развитие нервной системы позвоночных начинается с формирования нервной пластинки из дорзальной эктодермы. Нервная пластинка затем сворачивается, формируя нервную трубку и нервный гребень. Нейроны и глиальные клетки в ЦНС образуются в результате деления клеток-предшественников вентрикулярной зоны нервной трубки. Постмитотические нейроны мигрируют от вентрикулярной поверхности нервной трубки и образуют серое вещество НС взрослых позвоночных. В пределах каждой из областей в развивающейся нервной системе судьба клеток в значительной степени зависит от их расположения. Недавно появилась возможность установить механизмы развития мозга на молекулярном уровне, что позволило объяснить такие механизмы, как рострокаудальный и дорзовентральный паттерны развития, которые до этого были описаны только феноменологически, без каких либо идей о механизмах. Например, экспрессия серии гомеобоксных генов вдоль рострокаудальной оси способствует дифференцировке сегментов заднего мозга; дорзовентральный паттерн определяется отчасти градиентом протеина, известного как Sonic hedgehog.
Клетки нервного гребня образуют периферическую нервную систему. Фенотип, в который дифференцируются клетки нервного гребня, определяется сигналами, поступающими от соседних клеток. Таким образом, если клетки нервного гребня были пересажены в раннем возрасте, они будут дифференцироваться в соответствии со своим новым расположением.
Для установления синаптических контактов со своими мишенями, нейроны образуют аксоны, имеющие на концах конусы роста, которыми они исследуют окружающее пространство. Было идентифицировано два класса молекул, играющих важную роль в продвижении конусов роста: молекулы клеточной адгезии надсемейства иммуноглобулинов и молекулы адгезии внеклеточного матрикса. Навигация конуса роста контролируется при помощи аттрактантов и репеллентов, действующих на коротких и длинных дистанциях. Хемоаттрактанты управляют ростом аксона либо до конечной, либо до промежуточной цели, например до клетки-ориентира (guidepost cell).
Филогенез нервной системы
... - этап нервной жизни. 1. Филогенез Филогенез - это процесс исторического развития вида. Филогенез нервной системы - история. Формирования и совершенствования ее структур. Филогенез нервной системы в кратких чертах сводится к следующему. У простейших одноклеточных организмов (амеба) нервной системы еще ...
Хеморепелленты не допускают вторжения аксонов в определенные «неблагоприятные» зоны. Проекции аксонов во время развития зачастую более сильно развиты, чем таковые у взрослых, у которых осуществляется усечение связей на основе механизмов, связанных с активностью и трофическими факторами.
Функциональные синаптические контакты формируются быстро, однако сначала они не имеют характерной специализации, свойственной синапсам взрослых. Только по прошествии нескольких недель синапсы созревают и приобретают свойства синапсов взрослых.
Характерной чертой развития центральной нервной системы всех позвоночных является первоначальное образование избыточного количества нейронов, часть из которых в дальнейшем погибает. Гибель нейронов регулируется при помощи конкуренции за трофические факторы. Фактор роста нерва является представителем семейства протеинов (нейротрофинов, neurotrophins), которые способны поддерживать активность определенных популяций нейронов.
Упорядоченность связей, образованных нервными клетками между собой, а также между ними и другими тканями на периферии является одним из условий для нормального функционирования нервной системы. Для создания такой точной нервной архитектуры во время развития необходимо, чтобы сформировались нейроны определенных типов в необходимом количестве и в строго определенных местах нервной системы, которые затем должны образовать связи со своими уникальными клетками-мишенями. Например, чтобы работал рефлекс растяжения, необходимо, чтобы афферентный сенсорный нейрон типа Ia спинномозгового ганглия послал свой аксон к определенному месту в области мышечного веретена, а другой аксон, идущий в ЦНС, должен образовать синаптический контакт исключительно с тем мотонейроном, который иннервирует мышцу, где находится это мышечное веретено. Другие веточки этого центрального аксона заканчиваются на спинальных интернейронах или уходят в дорзальные столбы для иннервации клеток, расположенных в ядрах спинного мозга. Кроме того, количество сенсорных и моторных нейронов должно находиться в определенном соотношении в зависимости от размеров мышцы и количества мышечных веретен, которые эта мышца содержит.
Большой спектр вопросов возникает при рассмотрении данного примера. Каким образом клетки дифференцируются в нейроны или клетки глии? По каким меткам следует нейрон, мигрируя к своему месторасположению? Какие клеточные механизмы позволяют нейрону направлять свой аксон строго к определенной цели, одной из мириада возможных, чтобы образовать на ней синапс? В поиске ответов на эти вопросы мы вспоминаем, что в нервной системе имеется 1010-1012 нервных клеток, причудливо связанных друг с другом, при этом количество генов, которые могут определять эту структуру, составляет всего 105 или около того. Более того, ЦНС демонстрирует большую гибкость во время критических периодов развития и даже у взрослых, у которых образуются новые синапсы, а старые либо изменяют свои свойства, либо просто исчезают, — и все это как результат изменения активности клеток.
Спектр проблем, относящихся к развитию, образованию синапсов, нервной специфичности и изменению свойств связей, настолько широк, что дать их полный обзор не представляется возможным. Многие аспекты этого уже подробно освещены в других руководствах. В этой главе мы осуществим короткий экскурс в нейроэмбриологию и опишем отдельные экспериментальные подходы к вопросам развития нервной системы. Темы будут освещаться нами в том порядке, как происходит процесс развития НС: начиная с нейроэктодермы и раннего морфогенеза нервной системы, затем мы рассмотрим региональную спецификацию нервной ткани и факторы, которые определяют идентификацию отдельных нейронов и клеток глии, и в конце мы поговорим о механизмах роста аксонов, иннервации ими определенной цели, образовании синапсов, а также о роли факторов роста и конкуренции в окончательном формировании нервной системы.
Становление и развитие системы профессионального образования в России
... развития профессионального образования в Российской империи в XIX – начале ХХ вв. Изучить особенности развития профессионального образования в Советском Союзе. Объект изучения – история становления и развития системы профессионального образования в России. Предмет изучения – особенности и аспекты истории становления и развития системы профессионального образования ...
Терминология
Основная терминологическая проблема возникает при изучении развития на молекулярном уровне. В последние годы описывается все больше и больше молекул, для которых известен механизм их действия. Сюда относятся также белки и гены, важные для выживания клетки, ее роста и дифференцировки, для формирования аксона и его навигации, для образования синапса и управления его свойствами. Для того, чтобы еще больше усложнить ситуацию, большинство этих белков и генов получили своеобразные имена, связанные с историей их открытия или благодаря выдумке их первооткрывателей в лабо должен был познакомиться с таким чужеродным понятием, как «замедленное выпрямление» (delayed rectification) из нашей дискуссии о передаче сигнала, сейчас вынужден привыкать к серии новых необычных названий, таких как «Sonic hedgehog» («звучащий ежик») и эфрин («Ephrin»), a также к бесконечному числу сокращений, типа «N-CAM», «BDNF» и «Elf-2», ни одно из которых непосредственно не говорит о функции того или иного агента. Мы постарались представить здесь минимальное количество генов и белков и, где возможно, использовать их полные имена.
Генетические подходы к пониманию процесса развития
Основные достижения последних лет привели к значительному скачку в нашем понимании молекулярных механизмов тех процессов и явлений, которые ранее были только констатированы. Первым достижением является разработка новых молекулярных биотехнологий для контроля и управления экспрессией генов. Вторым является открытие механизмов и молекул, которые управляют развитием нервной системы удивительно сходным образом у различных животных. Например, как мы уже упоминали в главе 1, гены, которые управляют развитием глаза у развивающегося цыпленка, мышонка или человека очень сходны с генами, определяющими формирование глаза дрозофилы. Таким образом, гены, определяющие развитие дрозофилы, грибов-дрожжей и нематод (C.elegans), часто имеют гомологов среди генов, определяющих развитие позвоночных. Третьим достижением является использование полосатой коралловой рыбы (zebra fish, официальное русское название «полосатый данио», Brachydanio rerio), впервые предложенной Страйзингером, как особенно удачного объекта для изучения развития позвоночных. Эмбрион полосатого данио прозрачен, что позволяет непосредственно наблюдать за отдельными клетками во время эмбриогенеза, который у данио происходит довольно быстро. Наиболее важным является то, что были разработаны приемы, при помощи которых у данио могут быть вызваны, контролированы и поддержаны на определенном уровне направленные мутации, что открывает дорогу к обнаружению важных генов позвоночных, которые могут иметь гомологов у некоторых видов более просто устроенных беспозвоночных.
Особенности распространения возбуждения в центральной нервной системе
... нервных центров. Нервный центр - совокупность нервных клеток, более или менее строго локализованная в нервной системе и непременно участвующая в осуществлении рефлекса или другого вида нервной деятельности. Основными свойствами передачи возбуждения являются: одностороннее проведение возбуждения; задержка проведения возбуждения; суммация возбуждений; ...
Развитие нервной системы в раннем периоде
В раннем периоде морфогенеза позвоночных область гаструлы, из которой в дальнейшем формируется нервная система, представляет собой простую полоску эктодермы (рис. 1).
Клетки этой полоски находятся под влиянием факторов роста (включая два белка из семейства костных морфогенетических белков, ВМР-2 и ВМР-4), которые подавляют нервную дифференцировку и приводят к развитию эпидермальной ткани. Затем из особого «организационного» региона гаструлы, названного Шпемановским организационным центром (Spemann organizer, в яйцах амфибий) или Гензеновским узелком (Hensen’s node, у эмбрионов цыпленка и млекопитающих), происходит выделение свободно диффундирующих белков, которые блокируют действие этих факторов роста. Эти белки (известные как фоллистатин (follistatin), ноггин (noggin) и хордин (chordin)) активируют сигнальные каскады в клетках, расположенных рядом с организационным центром, что приводит к дифференцировке клеток в нейроны и формиро ванию нервной пластинки. Нервная пластинка представляет собой полоски продолговатых нейроэктодермальных клеток, из которых в дальнейшем будет сформирована нервная система.
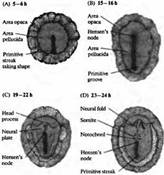
Рис. 1. Ранний морфогенез в эмбрионе позвоночного. Вид сзади на развивающийся эмбрион цыпленка в первый день жизни. (А) 5-6 ч.: образование и удлинение первичной полоски (В) 15-16 ч.: образование первичного желобка и Гензеновского узелка. (С) 19-22 ч.: образование головного отростка и нервной пластинки. (D) 23-24 ч.: образование нервного валика, хорды и мезодермальных сомитов.
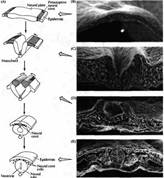
Рис. 2. Образование нервной трубки в эмбрионе цыпленка. (А) Диаграмма нейруляции (В-Е) Микрофотограммы образования нервной трубки, сделанные при помощи сканирующего электронного микроскопа. (В) Нервная пластинка, образованная продолговатыми клетками в дорзальной области эктодермы. (С) Нервный желобок, образованный продолговатыми неирозпителиальными клетками и окруженный мезенхимальными клетками. (D) Нервные валики, покрытые уплощенными зпидермальными клетками. (Е) Нервная трубка, покрытая эпидермисом с расположенными по обеим сторонам сомитами и хордой внизу.
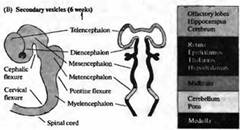
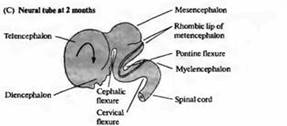
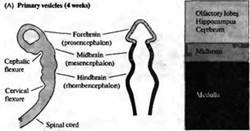
Рис. 3. Ранний период развития головного мозга человека. Вид сбоку на развивающийся мозг и схематический горизонтальный срез через пузыри. (А) На 4 неделе развития ЦНС человека состоит из трех первичных пузырей. (В) К 6 неделе развития можно различить пять вторичных пузырей. (С) В возрасте 2 месяцев серии складок, сужений и утолщений образуют различные области мозга. Дальнейшее развитие характеризуется преимущественным ростом конечного мозга в форме буквы «С» (показано стрелками).
Эволюция нервной системы
... развития. По-видимому, хордовые животные происходят от низших беспозвоночных животных, нервная система которых была организована диффузно. ЦНС хордовых имеет вид сплошной нервной трубки (трубчатая нервная система), которая у позвоночных животных образует в переднем конце ...
Нервная пластинка затем утолщается по своему краю, края ее приподнимаются, образуя нервные валики, которые сливаются друг с другом по средней линии, образуя полую нервную трубку (рис. 2).
Процесс, благодаря которому осуществляется формирование нервной пластинки и превращение ее в нервную трубку, называется нейруляцией Некоторые из клеток, расположенные по краям нервных валиков, остаются между нервной трубкой и вышележащей эктодермой. Эти клетки формируют нервный гребень. Клетки нервного гребня мигрируют в направлении от нервной трубки и образуют большое разнообразие периферических тканей, в том числе нейроны и клетки-спутники в сенсорной, симпатической и парасимпатической нервной системе, клетки надпочечника, пигментные клетки эпидермиса, кости и соединительные ткани в области головы.
По мере развития в передней (головной или ростральной) части нервной трубки формируется серия утолщений, сужений и изгибов, образующих отдельные анатомические области мозга (рис. 3).
Каудальная часть нервной трубки остается относительно простой по строению, сохраняя вид полой трубки, и в дальнейшем на ее основе формируется спинной мозг.
Образование предшественников нервных клеток и глии
Стенка нервной трубки первоначально состоит из одного слоя быстро делящихся клеток. Каждая клетка простирается от полостного, вентрикулярного края к наружной, пиальной поверхности. По мере того, как каждая клетка проходит свой клеточный цикл, ее ядро мигрирует вверх и вниз от вентрикулярной (желудочковой) к пиальной поверхности (рис. 4А).
Синтез ДНК происходит в то время, когда ядро расположено у пиальной поверхности; во время деления клетки (cytokinesis) ядро лежит у вентрикулярной поверхности и на некоторое время клетка теряет связь с пиальной поверхностью. После деления одна или обе дочерние клетки могут потерять контакт с вентрикулярной поверхностью и мигрировать вверх. Этот момент является переломным: они превращаются либо в нейроны, либо в клетки глии. Если они мигрируют в направлении от вентрикулярной зоны, большинству этих клеток суждено стать нейронами в постмитотической фазе (то есть они уже никогда не смогут делиться. Предшественники глиальных клеток, с другой стороны, могут делиться даже после достижения своего окончательного месторасположения.
По мере того, как все больше и больше формируется постмитотических клеток, нервная трубка утолщается и приобретает трехслойное строение: наиболее глубоко расположена вентрнкулярная зона (где продолжается деление клеток), затем средняя зона в виде мантии (плащевая зона), где находятся тела мигрирующих нейронов, а также поверхностный краевой (маргинальный) слой, состоящий из отростков более глубоко расположенных клеток (рис. 4В).
Такая трехслойная структура сохраняется в спинном и продолговатом мозге (рис. 4С).
В других областях, таких как кора больших полушарий и мозжечок, некоторые нейроны мигрируют в краевую зону и формируют корковую пластинку, которая затем созревает и формирует взрослую кору.
Миграция нейронов в ЦНС
Во многих областях развивающегося мозга, например в коре полушарий и мозжечке, миграция нейронов зависит от радиально расположенных глиальных клеток . Эти клетки поддерживают контакты с вентрикулярной и пиальной поверхностями нервной трубки. По мере утолщения нервной трубки из-за продолжающегося деления клеток в области вентрикулярного слоя и накопления нейронов в промежуточной зоне мантии и кортикальной пластинки, радиальные клетки глии значительно удлиняются. При помощи детального изучения развития мозжечка и коры методами световой и электронной микроскопии Ракич с коллегами показали, что нейроны движутся по «этажерке» из радиально расположенных клеток глии для того, чтобы достичь своего правильного расположения в коре. Наблюдения за мутантными мышами и опыты на клетках, выращенных в культуре подтвердили данное наблюдении о миграции нейронов. Сейчас начинают описываться белки, управляющие подобной миграцией нейронов. Они представляют собой нейрональный гликопротеин, известный как астротактин (astrotactin) и изоформы семейства рецепторов интегрина к молекулам адгезии экстрацеллюлярного матрикса (о чем мы поговорим ниже).
Общая характеристика нервная ткани
... функциональным состоянием клетки. Цитоплазма всех нервных клеток богата рибосомами, которые, как и в клетках других тканей, представлены гранулами диаметром 150-350 Ǻ. В нейробластах рибосомы распределяются в матриксе свободно по одиночке или ...
Нейроны способны мигрировать также и в направлении тех областей нервной системы, где нет радиально расположенных глиальных клеток. Одна интересная популяция нейронов, экспрессируюших гормоны гонадотропина (gonadotropin-releasing hormone (GnRH)), мигрирует с периферии в ЦНС. GnRH клетки проходят расстояние около 2 мм, мигрируя из области обонятельной ямки, эктодермальное производное (плакода) которой также образует эпителий носа, в область гипоталамуса по пути предварительно спроецированного туда аксона.
Белки адгезии внеклеточного матрикса и миграция клеток нервного гребня
В периферической нервной системе клетки нервного гребня не мигрируют по путям предварительно спроецированного аксона или вдоль структур глиальных клеток. Их миграция управляется механизмами привлечения и отталкивания при контакте с поверхностью клеток и компонентами внеклеточного матрикса. Два белка адгезии внеклеточного матрикса, ламинин (laminin) и фибронектин (fibronectin), концентрируются вдоль путей миграции клеток нервного гребня у эмбриона . Агенты, которые угнетают взаимодействие между рецепторами интегрина и компонентами внеклеточного матрикса, блокируют перемещение in vitro клеток нервного гребня вдоль поверхности, покрытой молекулами адгезии внеклеточного матрикса, а также миграцию in vivo клеток нервного гребня. Другой белок внеклеточного матрикса, Ф-спондин (F-spondin), экспрессируется в областях, граничащих с маршрутами миграции клеток нервного гребня. Он ингибирует перемещение клеток гребня, таким образом направляя их на правильный путь
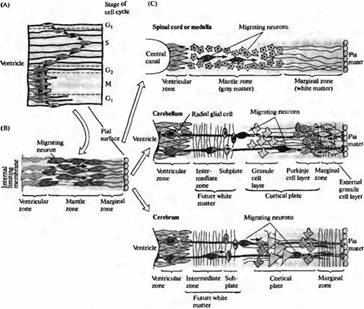
Рис. 4. Дифференцировка клеток стенок нервной трубки. (А) Расположение ядер в клетках в первичной нервной трубке в зависимости от стадии клеточного цикла. (В) Клетки, становясь постмитотическими, мигрируют прочь от вентрикулярной зоны и образуют плащевую зону. Их отростки направляются в краевую зону. (С) Трехслойная организация сохраняется в спинном мозге. В мозжечке и головном мозге нейроны мигрируют в краевую зону и образуют там многослойную кору.
Литература
[Электронный ресурс]//URL: https://psychoexpert.ru/referat/razvitie-nervnoy-sistemyi/
1. Arshavsky, Y. I., Deliagina, Т.О., and Orlovsky, G. N. 1997. Pattern generation. Cuir. О/ни. Neuro-biol. 7: 781-789.
Строение и функции нервной ткани
... обеспечивает неравновесное распределение указанных ионов между клеткой и внеклеточной средой, что лежит в основе формирования электрического заряда клетки. Механизм активного транспорта ионов поддерживает ионную ... в частности, усиливать ее эффективность. Другие виды классификаций нейронов. Нервные клетки разных отделов нервной системы могут быть активными вне воздействия, т. е. обладают ...
2. Asanuma, H. 1989. The Motor Cortex. Raven, New York.
3. Georgopoulos, A. P. 1994. New concepts in generation of movement. Neuron 13: 257-268.
4. Getting, P. A. 1989. Emerging principles governing the operation of neural networks. Anna. Rev. Neurosci. 12: 185-204.
5. Kiehn, O., et al. (eds.) 1998. Neuronal Mechanisms for Generating Locomotor Activity (Annals of the New York Academy of Sciences,