
Способность отвечать возбуждением на действие раздражителя является одним из наиболее характерных свойств живых клеток. Поэтому изучению проблемы возбудимости всегда уделялось большое внимание. Наблюдающийся в настоящее время стремительный рост наших знаний в этой области связан с широким использованием в физиологической практике новых, весьма тонких и точных приемов исследования, позволяющих раздражать одиночные возбудимые образования, регистрировать их электрическую активность, фиксировать на заданном уровне мембранный потенциал, непосредственно измерять ионные потоки через мембрану.
Особенно выдающихся успехов за последние десятилетия добились исследователи, изучающие механизмы генерации биопотенциалов в нервных волокнах и клетках. Вершиной этих достижений, несомненно, являются исследования английских ученых Hodgkin и Huxley, которые развили выдвинутую еще в начале этого столетия концепцию Bernstein и сформулировали современную мембранную теорию возбуждения. Теория Hodgkin–Huxley получила всеобщее признание, и ее авторы были удостоены в 1964 г. Нобелевской премии.
1. Понятие о биоэлектрических явлениях
Первые данные о существовании биоэлектрических явлений («животное электричество») были получены в третьей четверти XVIII в. при изучении природы электрического разряда, наносимого некоторыми рыбами при защите и нападении. Многолетний научный спор (1791 — 1797) между физиологом Л. Гальвани и физиком А. Вольта о природе «животного электричества» завершился двумя крупными открытиями: были установлены факты, свидетельствующие о наличии электрических потенциалов в нервной и мышечной тканях, и открыт новый способ получения электрического тока при помощи разнородных металлов – создан гальванический элемент. Однако первые прямые измерения потенциалов в живых тканях стали возможны только после изобретения гальванометров. Систематическое исследование потенциалов в мышцах и нервах в состояния покоя и возбуждения было начато Дюбуа-Реймоном (1848).
Дальнейшие успехи в изучении биоэлектрических явлений были тесно связаны с усовершенствованием техники регистрации быстрых колебаний электрического потенциала и методов их отведения от одиночных возбудимых клеток. С помощью внутриклеточных микроэлектродов удалось произвести прямую регистрацию электрических потенциалов клеточных мембран. Успехи электроники позволили разработать методы изучения ионных токов, протекающих через мембрану при изменениях мембранного потенциала или при действии на мембранные рецепторы биологически активных соединений. В последнее время разработан метод, позволяющий регистрировать ионные токи, протекающие через одиночные ионные каналы.
Особенности распространения возбуждения в центральной нервной системе
... облегчение проведения иррадиация торможение возбуждения Одностороннее проведение возбуждения в центральной нервной системе обусловлено наличием в нервных центрах синапсов, в которых передача возбуждения возможна только в одном направлении - от нервного окончания, выделяющего медиатор, к постсинаптической мембране. Задержка проведения возбуждения в нервных центрах ...
Итак, биоэлектрические явления в тканях – это разность потенциалов, которая возникает в тканях в процессе нормальной жизнедеятельности. Эти явления можно регистрировать, использую трансмембранный способ регистрации.
При таком способе регистрируются:
- потенциал покоя или мембранный потенциал;
- потенциал действия.
2 . Понятие о возбудимых тканях
Все клетки и ткани живого организма под действием раздражителей переходят из состояния относительного физиологического покоя в состояние активности (возбуждения).
Наибольшая степень активности наблюдается в нервной и мышечной ткани.
Главными свойствами возбудимых тканей являются: а) возбудимость; б) проводимость; в) рефрактерность и лабильность, которые связаны с одним из самых общих свойств живого – раздражимостью.
раздражителями
По природе раздражители бывают: механические, химические, электрические, температурные.
По биологическому признаку раздражители делятся на: 1) адекватные, которые воспринимаются соответствующими специализированными рецепторами (например, рецепторами глаза – свет, уха – звук, кожи – боль, температура, прикосновение, давление, вибрация); 2) неадекватные, к которым специализированные рецепторы не приспособлены, но воспринимают их при чрезмерной силе и длительности (например, удар – глаз — свет).
нервный импульс
Основные физиологические свойства нервной ткани (возбудимость, проводимость, рефрактерность и лабильность) характеризуют функциональное состояние нервной системы человека, определяют его психические процессы.
Возбудимость –
порог возбуждения
подпороговым
Разность потенциалов в покое между внутренним
мембранным потенциалом покоя
потенциалом действия
Следовательно возбуждение нервной клетки связано с изменением обмена веществ и сопровождается появлением электрических потенциалов (нервных импульсов).
Проводимость
пространственного взаимодействия
Рефрактерность
Такая динамика возбудимости обусловлена процессами изменения и восстановления равновесия ионов на мембране клетки.
Профессор Н.Е. Введенский исследовал особенности этих процессов и установил, что возбудимые ткани могут отвечать разным числом потенциалов действия на определенную частоту раздражений. Он назвал это явление лабильностью (функциональной подвижностью).
Лабильность
3. Мембранный потенциал покоя
мембранный потенциал
|
|
|||||
|
|
|||||
|
|
|||||
| |
|
|||||
|
||||||
|
||||||
Рис.1 Внутри- и внеклеточные концентрации ионов для клетки теплокровного животного, ммоль/л.
мембранным потенциалом покоя (МПП).
калиевым равновесным потенциалом
![]() =
= ![]() (ф.1),
(ф.1),
где R –газовая постоянаая, T – абсолютная температура, Z – валентность иона и F – постоянная Фарадея. Для калия эта величина составляет –97мВ.
4. Мембранный потенциал действия
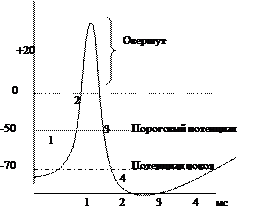
Если на нерв или мышцу нанести раздражение выше порога возбуждения, то МПП нерва или мышцы быстро уменьшается и на короткий промежуток времени (миллисекунда) произойдет перезарядка мембраны: ее внутренняя сторона станет заряженной положительно относительно наружной. Это кратковременное изменение МПП, происходящее при возбуждении клетки, которое на экране осциллографа имеет форму одиночного пика, называется мембранным потенциалом действия (МПД) (рис.2).
МПД в нервной и мышечной тканях возникает при снижении абсолютной величины МПП (деполяризации мембраны) до некоторого критического значения, называемого порогом генерации МПД. В гигантских нервных волокнах кальмара МПД равен –60 мВ. При деполяризации мембраны до –45 мВ (порог генерации МПД) возникает МПД.
Во время возникновения МПД в аксоне кальмара сопротивление мембраны уменьшается в 25 раз, с 1000 до 40 Ом/см 2 , тогда как электрическая емкость не изменяется. Указанное снижение сопротивление мембраны обусловлено увеличением ионной проницаемости мембраны при возбуждении.
реверсия заряда.
Причиной таких изменений мембранного потенциала является увеличение проницаемости мембраны для ионов натрия. Это происходит благодаря открытию натриевого канала. Натриевый канал представляет собой гликопротеид. Этот белок обладает способностью изменять свою конформацию при изменении величины мембранного потенциала.
ворот канала
Деполяризация мембраны, вызываемая раздражающим стимулом, например, электрическим током, открывает m – ворота натриевых каналов и обеспечивает появления направленного внутрь потока положительных зарядов – ионов натрия. Это ведет к дальнейшей деполяризации мембраны, что в свою очередь, увеличивает число открытых натриевых каналов и следовательно, повышает натриевую проницаемость мембраны. Возникает «регенеративная» деполяризация мембраны, в результате которой потенциал внутренней стороны мембраны стремится достичь величины натриевого равновесного потенциала.
Причиной прекращения роста МПД и реполяризации мембраны клетки является: а) увеличение деполяризации мембраны, т.е. когда мембранный потенциал (Е м ) → ЕNа , в результате чего снижается электрохимический градиент для ионов натрия, равный Ем — ЕNа. ( уменьшается сила, «толкающая» натрий внутрь клетки); б) деполяризация мембраны порождает процесс инактивации натриевых каналов (закрывание h- ворот), который тормозит рост натриевой проницаемости мембраны и ведет к ее снижению; в) деполяризация мембраны увеличивает ее проницаемость для ионов калия. Процессы инактивации натриевых каналов может быть блокирован токсинами и фармакологическими препаратами (местными анестетиками).
Выходящий калиевый ток стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала.
Снижение электрохимического потенциала для ионов натрия и инактивации натриевых каналов уменьшает величину входящего натриевого тока. В определенный момент времени величина входящего тока натрия сравнивается с возросшим выходящим током – рост МПД прекращается. Когда суммарный выходящий ток превышает входящий, начинается реполяризация мембраны, которая также имеет регенеративный характер. Начавшаяся реполяризация ведет к закрыванию активационных ворот (m), что уменьшает натриевую проницаемость мембраны, ускоряет реполяризацию, а последняя увеличивает число закрытых каналов и т.д.
рефрактерностью.
 |
 |
||
|
|||
|
|||
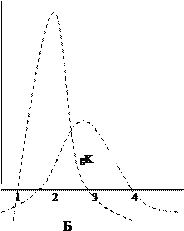
Рис. 2. Потенциал действия
А . График развития потенциала действия. Б . Изменения проницаемости мембраны в процессе развития потенциала действия.
1. – фаза медленной деполяризации; 2 . – фаза быстрой деполяризации; 3 .- фаза реполяризации; 4 .- фаза гиперполяризации.
4.1. Распространение потенциала действия
сальтаторное проведение
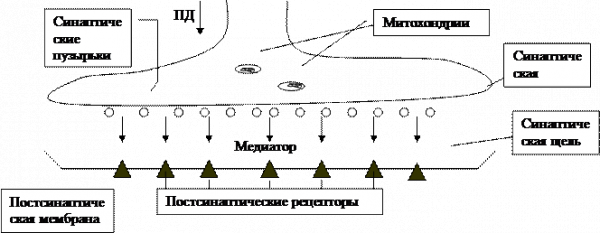
4.2. Межклеточная передача возбуждения
химические
 |
||
|
||
моноамины
Поступивший в синаптическую щель медиатор взаимодействует с рецептором постсинаптической мембраны, вызывая изменение ее проводимости. Механизм этого взаимодействия описан в рамках теории молекулярной рецепции – раздела биокинетики. Несколько слов об истории вопроса. В начале XX-го века П.Эрлих и И.И.Мечников сформулировали теорию боковых цепей (Нобелевская премия, 1908).
Согласно этой теории на клеточной поверхности имеются особые химические соединения, которые образуют «боковые цепи». Химические соединения способны взаимодействовать с этими боковыми цепями. В результате взаимодействия происходит клеточное распознавание веществ и формирование клеточного ответа. П.Эрлих назвал эти боковые цепи рецепторами (от латинского recipere – получать, узнавать).
Он сформулировал основной постулат рецепторного механизма действия химических соединений: “вещество не действует, если не фиксировано” (“corpora non agun nisi fixata”).
Первое количественное описание величины ответа клетки в зависимости от концентрации добавленного химического соединения было предпринято А. Кларком в 1926г. Аналогично тому, как это сделано в ферментативной кинетике, Кларк предположил, что химические вещества находятся в избытке по отношению к рецепторам. Кроме того, он предположил, что взаимодействие химических веществ с рецепторами обратимо. В настоящее время химические вещества, способные взаимодействовать с рецепторами, принято называть лигандами ( от латинского ligo – связываю).
Такое взаимодействие лигандов с рецепторами, согласно Кларку записывается так:
![]() (ф.2),
(ф.2),
где L –лиганд; R – рецептор; LR – комплекс лиганда с рецептором или лиганд-рецепторный комплекс; ![]() и
и ![]() — константы скоростей образования и распада лиганд-рецепторных комплексов.
— константы скоростей образования и распада лиганд-рецепторных комплексов.
Согласно современным представлениям, большинство биологически активных веществ и лекарств оказывает воздействие на организм, регулируя функции клеток. При этом многие вещества не проникают в клетку, а оказывают свое действие опосредованно, взаимодействуя на клеточной мембране со специфическими молекулами, называемыми рецепторами. Рецепторы способны специфически распознать биологически активное вещество, обратимо или необратимо взаимодействовать с ним и передавать специфический сигнал об этом взаимодействии в клетку. Под рецепторами понимают макромолекулы, расположенные на цитоплазматической мембране клеток или внутриклеточно, способные специфически взаимодействовать с ограниченным набором биологически активных веществ (лигандов) и трансформировать сигнал об этом взаимодействии в специфический клеточный ответ. Все лиганды делятся на агонисты и антогонисты . Агонистами называют лиганды, которые, связываясь с рецепторами данного типа, активно вызывают клеточный ответ . В синапсе в качестве агонистов могут выступать как медиаторы, так и другие вещества. Это обусловлено тем, что специфичность мембранных рецепторов не абсолютна. Так, в холинергическом синапсе в качестве агонистов могут выступать карбамилхолин и суберилдихолин. Другие вещества, также связывающиеся с рецепторами медиаторов, но не столь эффективно изменяющие мембранную проницаемость, называются частичными агонистами . Антагонистами называются лиганды, которые связываются с рецепторами и не вызывают активного клеточного ответа . Антагонисты препятствуют связыванию агонистов с рецепторами, угнетая клеточные функции. Связывание их может быть обратимым: спустя некоторое время антагонист отделяется от рецептора. Такие вещества называют конкурентными антагонистами, так как они конкурируют с медиаторами и их антагонистами за участки связывания. Хорошо известный конкурентный антагонист ацетилхолина в нервно-мышечном синапсе – яд кураре (d-тубокурарин).
По мере повышения концентрации он блокирует все больше рецепторов, и эффект ацетилхолина ослабляется из-за уменьшения доступных мест связывания. Это приводит к уменьшению потенциала концевой пластинки и параличу мышцы.
С учетом всего вышесказанного формула Кларка будет выглядеть следующим образом:
![]() (ф.3),
(ф.3),
рецепторы – ионные каналы
1) аденилатциклаза – циклический аденозин – моно – фосфат (цАМФ);
2) гуанилатциклаза – циклический гуанозин – монофосфат (цГМФ);
3) фосфолипаза С — инозитол – три – фосфат (Ифз);
4) ионизированный кальций.
В значительной части клеток организма присутствуют или могут образовываться почти все из перечисленных выше вторичных посредников (исключением является цГМФ).
Это обуславливает различные взаимосвязи между вторичными посредниками: 1) равноправное участие, когда разные посредники необходимы для медиаторного эффекта; 2) один из посредников является основным, а другой лишь способствует реализации эффектов первого; 3) посредники действуют последовательно (например, инозитол – 3- фосфат обеспечивает освобождение кальция, диацилглиццерол облегчает взаимодействие кальция с протеинкиназой С); 4) посредники дублируют друг друга для обеспечения избыточности с целью надежности регуляции; 5) посредники являются антагонистами, т.е. один из них включает реакцию, а другой тормозит (например, в гладких мышцах сосудов инозитол–3–фосфат и кальций реализуют их сокращение, а цАМФ – расслабление).
рецепторами, сопряженными с ферментами
1) ![]() — белки. Оказывают стимулирующее влияние на аднилатциклазу, оказывает ингибирующее влияние на медленный тип кальциевых каналов.
— белки. Оказывают стимулирующее влияние на аднилатциклазу, оказывает ингибирующее влияние на медленный тип кальциевых каналов.
2) ![]() — белки. Оказывают ингибирующее воздействие на аденилатциклазу, активируют цАМФ-фосфодиэстеразу.
— белки. Оказывают ингибирующее воздействие на аденилатциклазу, активируют цАМФ-фосфодиэстеразу.
3) ![]() -белки. Их активация приводит к угнетению потенциал-зависимых кальциевых каналов и стимуляции калиевых каналов.
-белки. Их активация приводит к угнетению потенциал-зависимых кальциевых каналов и стимуляции калиевых каналов.
4) ![]() -белки. Вызывают активацию калиевых каналов и угнетение аденилатциклазы.
-белки. Вызывают активацию калиевых каналов и угнетение аденилатциклазы.
5) ![]() -белки. Приводят к активации фосфолипазы С.
-белки. Приводят к активации фосфолипазы С.
возбуждающий постсинаптический потенциал (ВПСП)
В качестве гистологического субстрата пресинаптического торможения удобнее всего рассматривать аксо-аксональные синапсы. В ходе проведенных различными учеными исследований было установлено, что снижение количества высвобождаемого медиатора в случае пресинаптического торможения связано с активацией именно этого вида синапсов. В механизме пресинаптического торможения большую роль играет деполяризация первичных афферентов (ДПА).
Активация аксо-аксонного синапса сопровождается деполяризацией постсинаптической области. Такую деполяризацию можно зарегистрировать внутриклеточно в первичных афферентах спинного мозга, откуда она и получила свое название. ДПА сопровождается уменьшением амплитуды пресинаптического потенциала в результате инактивации натриевых каналов. В конечном счете, может происходить полная блокада проведения в пресинаптических окончаниях, сопровождающаяся высвобождением очень небольшого количество медиатора.
После того, как медиатор диффундировал через синаптическую щель, дальнейшая его диффузия из этого узкого пространства идет довольно медленно и не способна заметно снизить установившуюся концентрацию.
Важнейшим, с точки зрения физиологии, вопросом является взаимодействие между синапсами. И действительно, первый же шаг от этого элементарного уровня взаимодействия двух клеток возбудимых тканей вверх по шкале размерностей, первый же шаг к пониманию целостной функциональной организации нервной системы требует рассмотрения этого взаимодействия.
Объединяясь между собой посредством синапсов, нейроны формируют изумительные по сложности системы, чьё функционирование и определяет, в конечном счетё, высшие функции центральной нервной системы. Здесь мы рассмотрим лишь наиболее простые из этих нейронных сетей. Эти элементарные нейронные цепи служат, например, для усиления слабых сигналов, уменьшения слишком интенсивной активности, выделения контрастов, поддержания ритмов или сохранения рабочего состояния нейронов путём регулировки их входов. Сама собой напрашивается аналогия с интегральными электрическими цепями – стандартными элементами, которые выполняют наиболее часто повторяющиеся операции, и могут быть включены в схемы самых разнообразных электронных приборов. Эти простейшие нейронные системы расположены во всех отделах головного и спинного мозга и характеризуются некоторым набором структурно функциональных феноменов, к числу которых относится конвергенция, дивергенция, временная и пространственная суммация (облегчение), различные виды торможения и реверберации.
дивергенция
конвергенцию
временно́го облегчения
Для пространственного облегчения
простые тормозные цепи
окружающим торможением
Гомонимными, Заключение
Термином «электрическая возбудимость» следует обозначать свойство клеточных мембран отвечать на сдвиги мембранного потенциала специфическими изменениями ионной проницаемости. По мнению большинства исследователей, электрическая возбудимость нервных и большинства мышечных волокон связана с существованием в плазматической мембране этих образований раздельных и, по-видимому, пространственно разобщенных путей – «каналов», по которым может происходить движение ионов Na + и K+ вдоль электрохимического градиента.
Потенциал покоя при данных градиентах ионов Na + , K+ и CI+ определяется в нервных волокнах в основном соотношением коэффициентов проницаемостей Р к /Р Na , каждый из которых в свою очередь является функцикй числа открытых натриевых и калиевых каналов.
Условием возникновения потенциала действия является повышение потенциала внутренней стороны мембраны до критической величины. При раздражении нервного волокна электрическим током эта критическая деполяризация осуществляется в результате суммирования пассивного – электротонического – изменения потенциала с активной подпороговой деполяризацией мембраны.
Мембранная теория одержала несомненную победу над другими гипотезами, пытавшимися объяснить природу биопотенциалов.
Литература
[Электронный ресурс]//URL: https://psychoexpert.ru/kursovaya/elektrohimicheskaya-priroda-nervnyih-impulsov/
1. Варфоломеев, С.Д. Биокинетика: Практический курс. / С.Д. Варфоломеев, К.Г. Гуревич. – М.: ФАИР-ПРЕСС, 1999. – 720с.
2. Коситский, Г.И. Физиология человека / Г.И. Коситский. – М.: «Медицина», 1985. – 544 с.
3. Основы физиологии человека. Учебник для высших учебных заведений: в 2-х томах. Т 1. / Под ред. Ткаченко Б.И. – СПб.: Международный фонд истории науки, 1994. – 560 с.
4. Проничев, И.В. Лекции по физиологии центральной нервной системы / И.В. Проничев. – У., 2003. – 156 с.
5. Психофизиология. Учебник для вузов. / Под ред. Ю.И. Александрова. – СПб.: Питер, 2001. – 496 с.
6. Физиология человека: в 3-х томах. Т.1. Пер с англ. / Под ред. Шмидта Р. и Тевса Г. – М.: Мир, 1996. – 323 с.
7. Ходоров, Б.И. Проблема возбудимости / Б.И. Ходоров. – СПб.: «Медицина», 1969. – 301 с.